
Transporte nos vegetais
As
primeiras espécies de plantas eram unicelulares, e todas tinham os seus locais
de consumo muito próximos uns dos outros, por isso o transporte de nutrientes
era um processo tranqüilo.
Com a evolução dos vegetais, surgiram às plantas
pluricelulares, e o transporte
de nutrientes foi se tornando cada vez mais complexo, uma vez que os órgãos
fotossíntetizadores foram ficando mais distantes das raízes. Logo, houve um
impedimento da troca de substâncias entre as folhas e a raiz.
Para suprir a necessidade de um mecanismo de
transporte de nutrientes, os vegetais desenvolveram os tecidos condutores.
Os tecidos condutores são: xilema e floema.
Estes tecidos são fundamentais para a subsistência das plantas. Eles são
capazes de realizar a condução de água e nutrientes, graças ás suas células que
são capazes de captar e transportar essas substâncias rapidamente de uma região
para a outra.
Quando a planta não possui esses tecidos
condutores, o transporte é realizado através da difusão. Nas
plantas sem tecidos condutores especializados, o movimento de solutos e água é
muito simples, usando a osmose e difusão, segundo os gradientes de
concentração. Apesar disso, mesmo nas briófitas já existem células com
algum grau de especialização no transporte de substâncias.
Todos
os vegetais traqueófitos apresentam um sistema duplo de transporte de
substâncias, muito eficaz e grande valor adaptativo para o meio
terrestre. É através deste sistema que se realiza o movimento de água e
solutos, orgânicos
Transporte de seiva bruta
Ao absorver sais do solo por transporte ativo, a raiz fica hipertônica e a água entra nas células por osmose. Essa
entrada de água com os sais gera a pressão de raiz, que empurra a seiva para
cima pelos vasos lenhosos. Em vasos lenhosos essa pressão não é forte o
suficiente para levar água até o topo.
Para que a planta realize uma boa
fotossíntese, estômatos das folhas devem abrir-se, o que leva a perda de água
por transpiração.com isso as células das folhas ficam mais concentradas e por
osmose absorvem água e sais minerais dos vasos lenhosos, mas próximos.
ESTÔMATOS E CONTROLE DA TRANSPIRAÇÃO
Os
estômatos são formados por duas células
clorofiladas em forma de rins, as células-guarda. A entrada de água altera a
forma dessas células e abre-se um espaço entre elas. Na perda de água as
células-guarda murcham e o ostíolo se fecha.
A
absorção de água pelas células-guarda resulta da entrada de íons de potássio
através da membrana por transporte ativo. Quando a célula acumula potássio, sua
pressão osmótica aumenta e, por osmose, ela absorve água das células vizinhas.
Se o transporte ativo é interrompido, o potássio acumulado sai por difusão, a pressão osmótica diminui e a célula perde água. Os
fatores que estimulam a entrada e a saída de íons potássio e, conseqüentemente,
a abertura e o fechamento dos estômatos são a luz, a concentração de gás
carbônico e o grau de hidratação da planta.
Normalmente
os estômatos estão abertos de dia e fechados de noite. A absorção de energia
luminosa estimula o transporte ativo de íons potássio, que se acumulam na
célula. Sem luz, não há transporte ativo de potássio, que sai da célula e
provoca o fechamento dos estômatos.
Quando as células do
parênquima clorofiliano perdem água, elas secretam acido
abscísico, hormônio que inibe o transporte ativo de íons de
potássio, fechando os estômatos. A baixa concentração de gás carbônico na folha
estimula o transporte ativo de íons potássio e a abertura dos estômatos.
O efeito do acido abscísico supera o
estimulo de abertura provocado pela luz e pela baixa concentração de gás
carbônico. Esse predomínio garante que, na iminência de uma desidratação os
estômatos se fechem. Com isso, a transpiração ocorre apenas pela cutícula e a
perda de água é muito pequena.
TRANPORTE
DE SEIVA ORGÂNICA
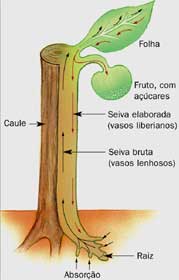 A matéria orgânica produzida nas folhas deve
ser distribuída para partes da planta que não fazem fotossíntese.
Esse transporte é realizado pelo floema.
A matéria orgânica produzida nas folhas deve
ser distribuída para partes da planta que não fazem fotossíntese.
Esse transporte é realizado pelo floema.
Nas células das folhas
forma-se a sacarose, que se difunde pelas células do parênquima clorofiliano
até o floema. Neste ela é absorvida por transporte ativo pelas
células-companheiras dos vasos
liberianos e passa para o interior da célula do vaso, ou seja, de uma pressão hidrostática, e não de
uma pressão osmótica.
Na outra extremidade do
floema, onde esta o órgão consumidor, o fluxo se faz no sentido contrário: as
células-companheiras bombeiam a sacarose do vaso liberiano para as células
órgão consumidor. Com a saída da sacarose, a pressão osmótica da célula do vaso
diminui e ela perde água para o órgão consumidor. Em conseqüência, a pressão
hidrostática nessa região diminui. Assim, a seiva move-se da região onde a pressão
hidrostática é mais alta para onde é menor.
 Essa teoria para o movimento da seiva
elaborada é conhecida como teoria do fluxo de pressão.
Essa teoria para o movimento da seiva
elaborada é conhecida como teoria do fluxo de pressão.
Os vasos liberianos estão
situados mais próximos á superfície do caule, na parte interna da casca. Se
fizermos um corte em anel na casca, o floema e a parte abaixo do corte deixam
de receber seiva orgânica, o que provocara a morte de suas células e da planta
por falta de nutrientes. Realizada pelo biólogo italiano Marcelo Malpighi, em
meados do século XVII, essa experiência demonstra o papel do floema no
transporte de seiva orgânica. Em homenagem ao cientista, experiência foi
chamada de anel de Malpighi.
Imagem extraída de:
http://www.portalsaofrancisco.com.br/alfa/anel-de-malpighi/padrao36.jpg
Referencias:
- Linhares,
Sergio/Gewandsznajder, Fernando, Biologia: volume único, pág.346, ano
2009, Ática.